
Sustainable Insect Management
Below here the text from our publication “Improving the efficacy of biological control by ecostacking” is cited , as an illustration of the principles involved in sustainable insect management:
Improving the efficacy of biological control by ecostacking. Chapter 1 in: Gao Y, Hokkanen HMT, Menzler-Hokkanen I (eds), Integrative Biological Control. Progress in Biological Control Volume 20, Springer-Nature, (Hokkanen HMT, Menzler-Hokkanen I, 2020). https://link.springer.com/chapter/10.1007%2F978-3-030-44838-7_1
——————————————————————————————————————————————-
Improving the efficacy of biological control by ecostacking
Heikki Hokkanen and Ingeborg Menzler-Hokkanen
Department of environmental and biological sciences, University of Eastern Finland, Yliopistonranta 1, FIN-70211 Kuopio, Finland
email: heikki.hokkanen@uef.fi
Abstract
Ecosystem service providers are fundamental to securing the long-term sustainability of our agricultural and food production systems. We will have to rely increasingly on ecosystem services to maintain and to increase agricultural production, and to make it sustainable in ecological, economical and social terms.
Biodiversity, or the lack of it, has been shown to affect the delivery of ecosystem services, including biological control, to crops. Diversity of ecosystem functions such as crop plant genetic variability, soil microbial assemblages, diversity of plant species in a field, and features of the agroecosystem landscape all play a role in determining the efficacy of the overall biological control in the target crop. The positive effects on enhancing biological control of pests, diseases and weeds, that can be obtainaed from the different levels of functional biodiversity, have not been utilised in a synergistic manner to optimize the level of biological control obtainable in a given environment.
We propose to make a full use of the benefits accruable from ecosystem services by conservation and stacking of functional biodiversity in our cropping systems, i.e. by “ecostacking” (Hokkanen 2017). Stacking involves combining, in an additive or synergistic manner, the beneficial services of functional biodiversity from all levels and types. This is a comprehensive approach, in which the different types of ecosystem service providers are fully integrated with the rest of the cropping system, including agronomic practices.
We illustrate here the principles of ecostacking with a case study, using the pollen beetle Brassicogethes aeneus, a key pest of oilseed rape, as our example. We provide evidence, and are confident that this problem pest can easily be maintained under complete biological control by combining several of the ecostacking component techniques into one management system.
Keywords: functional biodiversity, ecosystem services, integrated pest management, biological control, pollen beetle, Brassicogethes aeneus
Introduction
Developing cropping systems, which ensure that functional biodiversity including biological control agents can deliver their full potential for the benefit of agricultural production, is of utmost urgency. We have proposed that maximising the benefits obtainable from ecosystem services in a cropping system can best be ensured by a comprehensive approach called “ecostacking” (Hokkanen 2017, Hokkanen and Menzler-Hokkanen 2018a). Stacking implies combining the beneficial services of functional biodiversity from all levels and types in an additive or synergistic manner. The various ecosystem service providers and their needs must be fully integrated in the cropping system, including agronomic practices. The ecosystem service providers include many types of organisms and their functions such as beneficial arthropods, vertebrates, microbes, and plants. Insects, mites and spiders function as predators and parasitoids for the control of pests, but also as seed feeders and other specialist herbivores for weed control, and as pollinators of plants. Furthermore, organisms involved in decomposition, nutrient cycling, soil aeration and mixing, etc, need to be considered. For example, vertebrates such as small mammals, bats, and birds function as seed feeders and predators of pests, while microbes provide critical ecosystem services including pest, disease, and weed control. This can be delivered directly as components of “suppressive soils” (see Hokkanen and Menzler-Hokkanen 2018a), as colonisers of plants as endophytes, or as epiphytic microbial flora. That certain soils are suppressive to plant pathogens, has been known for a long time (e.g. Hornby 1983, Schlatter et al. 2017), but soils suppressive to insect pests has not been considered so far (but see Hokkanen and Menzler-Hokkanen, 2017, 2018b). How endo- and epiphytic micro-organisms on plants can steer arthropod–plant interactions is currently under intensive study, but applications based on microbe-mediated bottom-up effects in crop protection have not yet reached practical applications (Shikano et al. 2017). How plants can provide ecosystem services related to pest, disease and weed management, includes the crop plants themselves (e.g., via the beneficial effects based on genotypic diversity, and the ability to activate induced resistance mechanisms), companion crops (e.g., trap crops, intercrops, undersown crops), and plants providing nectar and pollen resources to beneficial arthropods.
In order to decide, which techniques can best be used to support the most essential biological control agents (BCA) and their functions in each specific cropping situation, it is necessary to (i) determine the role of the BCA under real field conditions in providing the biocontrol services; (ii) determine the most reliable and cost-effective sources of BCA either from the surrounding off-crop habitats, or from sources within the crop; (iii) investigate the possibilities of enhancing functional BCA by engaging all possible levels of biodiversity in the cropping system (ranging from molecular and genetic levels to microbial, vegetation, and landscape level), and stacking these to optimise the effect; and to (iv) ensure that the agricultural system as a whole is supporting these processes for example by replacing toxic pesticides with biocontrol products and biorational pesticides, and by ensuring that the pesticides and other techniques, which still need to be used, do not harm the BCA.
As an example, we outline next the principles of ecostacking for a concrete case, as an inspiration to adopt similar approaches for other situations, and to illustrate how a complete biological control of a problem pest could be achieved in annual cropping systems by combining multiple elements of ecostacking. Depending on the specific cropping context, the number of these elements necessary for full control may vary and should be studied and optimised locally.
Principles of ecostacking in the context of pollen beetle management
The pollen beetle Brassicogethes aeneus has been the most problematic pest of oilseed rape and turnip rape (“canola”) in Europe since decades. Locally and occasionally several other pest species may be more important, but the pollen beetle as a key pest has been more widespread, and a more constant threat to successful rapeseed production than any of the other species (e.g., Menzler-Hokkanen et al. 2006). Evolutionary adaptation to rapeseed growing practices, increasing the fitness of the pest, may have contributed to the pest status of the pollen beetle (Hokkanen 2000). The main control method against the pest has been, and continues to be, chemical control, mainly with synthetic pyrethroids over the last decades. Rapeseed fields are sprayed annually at least once, occasionally twice, and up to four times per season for pollen beetle control (Menzler-Hokkanen and Hokkanen 2018). Due to increasing resistance to synthetic pyrethroids in the pollen beetle (e.g., Tiilikainen and Hokkanen 2008), there is an urgent need to reduce the use of chemical pesticides, and to replace routine spraying with alternative control methods.
Landscape level processes
The pollen beetle overwinters as adult, and needs off-crop habitats for overwintering, selecting suitable sites within a range of several kilometers from the rapeseed fields (Müller 1941). The quality of the overwintering site undoubtedly affects the survival rate of the beetle, which can vary enormously. Hokkanen (1993) measured an overwintering survival rate of only 2-4% for a natural population in Finland, while later studies have found much higher survival rates (up to 20% in Northern Germany and 10-15% in Finland; Hokkanen, unpublished). Furthermore, the presence or absence of early flowering plants at the overwintering site, offering pollen food to the beetles after emergence from hibernation, is expected to greatly influence pollen beetle survival. The beetles become active after temperatures rise to about +5 °C or above, but cannot fly away from the hibernation site until the temperature reaches about +12 °C (Mauchline et al. 2017). In the absence of pollen food they face starvation, unless they are able to fly away to early flowering spring plants such as dandelions. The beetles are strong fliers (up to 15 km measured, Taimr et al. 1967) and are able to optimize habitat use for their benefit within the broad landscape. How the spatio-temporal dynamics involved in pollen beetle management is affected by agricultural landscapes, needs further studies and is presently not well understood.
In contrast, the key natural enemies – hymenopteran parasitoids – are relatively little affected by landscape features. Main parasitoids overwinter as pupae in the soil of the rapeseed field and emerge in the spring at the time of start of bud formation in the crop. Landscape affects their capacity to find and parasitize the pollen beetle larvae at least in two ways: (i) depending on crop rotation patterns, the next year’s rapeseed fields may be far away from the previous year’s fields. In that case the landscape will affect the ability of the parasitoids to find the new fields of rapeseed, and increases mortality in transit; (ii) availability of floral resources to provide nectar nutrition to the parasitoids searching for the target habitat and the host larvae in the new rapeseed fields (Hatt et al. 2018).
Landscape quality has been shown to significantly affect the level of pollen beetle parasitism in rapeseed, and consequently the damage caused by the pest: parasitism increases linearly as the proportion of uncultivated land within 1.5 km of the rapeseed field increases, and the amount of damaged buds decreases linearly at the same time (Thies and Tscharntke 1999).
Studies in Finland revealed the existence of regional level processes that synchronise host-parasitoid dynamics over large geographical areas (Hokkanen 2006). Data on annual percent parasitism of the pollen beetle larvae by Phradis morionellus (Ichneumoniidae) were collected in Finland for 11 years (1985-1995) from 35-70 different locations (13 regions) covering the total area of rapeseed growing in Finland. Correlation analysis of the time-series data for the 13 regions revealed that the parasitoid dynamics follow a surprising pattern, being in synchrony over several large regions, but varying widely between these larger regions (more than 100 km in diameter). It has been suggested (Hokkanen 2006) that only edaphic and climatic factors can explain such regional patterns, and indeed, the boundaries of relief pattern types, and those of biotic zones in Southern Finland are similar to the approximate boundaries for the parasitism dynamics (Hokkanen 2006).
In the rapeseed growing system, improving ecosystem service provision in the target crop by optimizing provision of off-crop resources, involves the provision of habitat, food, and alternative hosts for crop pollinators and natural enemies of crop pests. Providing early-flowering resources near the target crop will attract and maintain pollinators to the crop vicinity before the focal crop flowering period, enabling better pollination. Late-flowering resources support pollinators after crop flowering, enabling pollinators to finish the season cycle. Likewise, early flowering resources support natural enemies of crop pests when resources are scarce, helping to build-up natural enemy populations in the crop early enough to prevent pest outbreaks, and late-flowering resources provide food, habitat and alternative prey species after mass-flowering crops have finished blooming, therefore promoting better survival of natural enemies for the next season.
Role of field margins and trap crops as tools in ecostacking
Off-crop elements can facilitate benefits for ecostacking purposes. There is an obvious connection between field margins, and specifically sown trap and nursery crops (the “botanical triad”, Shrestha et al. 2019), to the landscape-level processes discussed above. In addition, these off-crop elements can improve pollination and pest control, provide habitat for farmland biodiversity, and maximize use of space to increase overall crop productivity. Trap crops (Hokkanen et al. 1986, Hokkanen 1991, Shelton and Badenes-Perez 2006) can be used directly for pest management, and can facilitate optimally timed delivery of emerging biocontrol technologies such as entomovectoring with insect pathogens (Hokkanen and Menzler-Hokkanen 2007, Hokkanen et al. 2015) and RNAi-products (Hokkanen and Menzler-Hokkanen 2018c). Timing in this context is critical, and trap cropping in the spring as well as in the autumn needs to be considered. Autumn trap crops may be easier to implement (e.g., less competing flowering plants) and can be superior for example in delivering next generation biopesticides based on RNAi (Hokkanen and Menzler-Hokkanen 2018c). In the spring, turnip rapeseed can sometimes effectively be used as a trap crop for protecting oilseed rape from pollen beetle attack (Hokkanen 1989).
In-field botanical diversity
Cropping systems based on designed ecological interventions such as mulches, or species or variety mixtures, have several advantages. These advantages include higher yield quality and stability, improved pest, weed and disease control, enhanced ecosystem services, and greater profitability (Balzan 2017). In general, variety mixtures can increase yield stability and suppress insect pests (Grettenberger and Tooker 2017).
Species or variety mixtures probably have never been tried or considered in the context of providing protection against the damage by the pollen beetle. Although there are distinct differences between pollen beetle preferences for different crucifer oilseed species (Ekbom and Borg 1996), this may not be useful for limiting pollen beetle damage in practice. Very little information is available concerning varietal preference (but see Herve et al. 2014, Herve and Cortesero 2016).
Possible impact on pest control in rapeseed of intercropping with various species of clover was studied in Finland (Perälä 1995). Strip intercropping with 50 cm broad strips of rapeseed alternating with 50 cm strips of four different species of clover, was compared with rapeseed monoculture. No differences were shown for the number of pollen beetle adults or larvae per plant, and no differences were observed in percent-parasitism by parasitic wasps, nor in the activity density of predatory ground beetles or spiders. The crop yields/ha (for the rapeseed strips in the intercrop-treatment) were equal, but the production of second generation pollen beetles/m2 was significantly higher (more than double) in the monocrop than in the intercrop system (Perälä 1995, Hokkanen 2005). This was explained by the higher predator pressure on pollen beetles during pupation and adult emergence in the intercrop system.
Hokkanen and Menzler-Hokkanen (2017, 2018a) argue for the ecological benefits of growing white clover as an undersown crop in spring rapeseed in Finland. Based on several years of practical field data (unpublished) it is clear that white clover as undercrop can effectively suppress weeds in the rapeseed growing system. White clover provides additional benefits for soil processes including nitrogen fixation and increases essential functional biodiversity in the system (see next section).
Soil biotic and abiotic properties
A key factor in determining the extent and impact of in-crop generation of biocontrol services is soil properties and management, along with soil cover management. Soil management and soil cover management are greatly affecting the soil and epigeial beneficial fauna (predatory mites, ground beetles, spiders, overwintering parasitoids, entomopathogenic nematodes, etc) and flora (entomopathogenic fungi and other microbes). At the landscape level, abiotic factors such as soil type and texture via their differential impact on parasitoid dynamics, are suspected as the main drivers of the regional synchronisation of patterns in pollen beetle parasitism (Hokkanen 2006).
Hokkanen and Menzler-Hokkanen (2018a) proposed to develop the concept of “insect pest suppressive soils” and provided data on the occurrence and importance of soil-borne insect pathogens in pest population suppression. Agricultural fields usually harbour only low numbers of beneficial insect antagonists such as entomopathogenic nematodes (EPN) and fungi (EPF), so that their role in pest population dynamics currently is negligible. Simple improvements in field and crop management, however, can quickly increase the numbers of EPN and EPF to levels that will lower the peak pest population levels (Hokkanen and Menzler-Hokkanen 2018a).
Concerning specifically the known pathogens of main oilseed Brassica pests, only entomopathogenic nematodes (EPN) and fungi (EPF) are widely distributed (Hokkanen et al., 2003). The EPF usually infecting pollen beetles are facultative pathogens, widely distributed in soils throughout the world, and occurring throughout the Brassica oilseed crops. Several studies have been published concerning their distribution in America, Australia and Europe (e.g., St Leger et al. 1992, Roberts and St Leger 2004). Studies often report a general relationship of these pathogens with the soil type, preferences of habitat type, and other abiotic and biotic factors affecting their abundance and role as insect antagonists (Vänninen 1996). However, their reported natural abundances in agricultural fields have always been low (e.g., Vänninen et al., 1989).
Most thoroughly studied EPF for potential control of oilseed Brassica pests include Beauveria bassiana and Metarhizium anisopliae. Both taxa have repeatedly been shown to kill effectively the adults (Fig. 1) and larvae of the pollen beetle (e.g., Butt et al. 1994; Husberg and Hokkanen 2001; Hokkanen et al., unpublished). In the field, treatments with EPF have not reduced the new generation pollen beetle numbers, neither via soil treatment nor via fungal sprays (Hokkanen 1993). However, soil treatments with EPF have caused dramatic indirect effects via increased overwintering mortality. Soil treatment with B. bassiana reduced overwintering survival of beetles developed under unlimited food resources by 50% (from 14% to 7% survival), compared with the untreated control.
and larvae of the pollen beetle (e.g., Butt et al. 1994; Husberg and Hokkanen 2001; Hokkanen et al., unpublished). In the field, treatments with EPF have not reduced the new generation pollen beetle numbers, neither via soil treatment nor via fungal sprays (Hokkanen 1993). However, soil treatments with EPF have caused dramatic indirect effects via increased overwintering mortality. Soil treatment with B. bassiana reduced overwintering survival of beetles developed under unlimited food resources by 50% (from 14% to 7% survival), compared with the untreated control.
The weight (fat reserves) of the beetles is a crucial factor explaining overwintering mortality of B. aeneus (Hokkanen 1993). Soil treatment with B. bassiana lowered the weight of pollen beetles developed under unlimited food resources by 16%. This alone is likely to be enough to result in the measured reduction in survival by 50%. However, beetles developing under normal conditions (collected from the wild) and under shortage of autumn food sources (pollen plants), were before overwintering 46% lighter than the experimental reference beetles. This lighter weight resulted in only 3% survival over the winter (Hokkanen 1993). If we assume that all rapeseed fields would contain effective levels of EPF either naturally or via augmentation, the pollen beetle populations would be expected to crash completely during the overwintering. Survival might be reduced from the measured 3% to maybe around 1%.
In contrast to the effect of EPF, direct mortality of pollen beetles in the field caused by entomopathogenic nematodes can be very high. Augmenting the field soil of rapeseed with Steinernema feltiae at the rates of 0.1-1.0 million infective juveniles (IJ) per m2 resulted in 95% reduction in the number of emerging pollen beetle adults (Hokkanen 2008). This shows a very high potential of the field soil to be suppressive to the pollen beetle, provided that adequate levels of antagonists such as EPF and EPN are present in the soil.
Plant associated microbes and their metabolites have been shown to have the potential to significantly alter the above-ground insect community. Plant associated microbes can affect virtually every type of plant-plant, plant-pathogen, plant-beneficial organism and plant-herbivore interactions (Pineda et al. 2017). A number of fungal and bacterial strains that occur naturally as part of the root microbiome, or indeed are used for biocontrol (e.g. Trichoderma spp., Bacillus spp., mycorrhizal fungi, Pochonia spp. and Paecilomyces spp.), have an indirect effect against pests via the plant, by activating defense response pathways. This induction can in some cases be more effective than the biocontrol agent itself and offers large but unexplored potential for ecostacking in the rapeseed cropping system.
The role of pesticides
Insecticide sprays to control peak population densities of the pollen beetle is routinely used in rapeseed cropping systems. In situations where the parasitoid occurrence and impact is low, as in winter oilseed rape in Germany, insecticide applications were not observed to affect significantly the parasitization of pollen beetle larvae by its ichneumonid parasitoids. This finding is regardless whether applied at the bud stage, at the beginning of flowering, or at full flowering (Brandes et al. 2018). In spring rapeseed, parasitoids often occur at much higher levels (e.g., Hokkanen et al. 1988, Veromann et al. 2009) and insecticide applications will affect parasitoids detrimentally if they take place close to flowering (Hokkanen et al. 1988). Spray window in Finland was determined to be before the accumulated day-degrees reach 320, normally at about mid-June. Insecticide treatments prior to that were considered to be safe to the parasitoids (Hokkanen et al. 1988).
An analysis of long-term patterns of pollen beetle parasitism in Finland revealed an interesting connection between the regularly occurring, severe crashes in percent-parasitism of the pollen beetle, and outbreaks of cereal aphids: parasitism levels always crash in years of cereal aphid outbreaks (Fig. 2; Hokkanen et al., unpublished).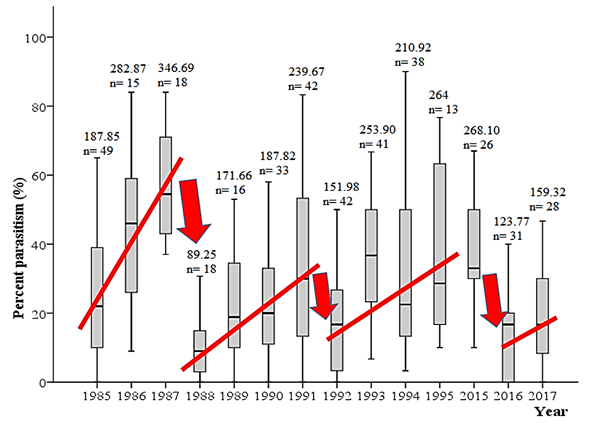 A strong negative correlation was found between pollen beetle parasitism level and the severity of cereal aphid attack, as well as between sales of the insecticide dimethoate (used to control aphids in cereals) and pollen beetle parasitism levels (Hokkanen et al. unpublished). It appears clear that in outbreak years of cereal aphids – about twice in a decade in Finland during the past several decades – the pollen beetle parasitoids are killed while the aphids are controlled with insecticide sprays. In crop rotations in Finland, spring oilseed crops are usually followed by spring cereals. Therefore, the pollen beetle parasitoids emerge in the following year from fields of spring cereals, usually exactly at the same time as when aphids colonise the fields and are controlled if needed (around mid-June till end of June). It would be best not to spray insecticides in those fields, where rapeseed was grown in the previous year, in order to maximize the biocontrol of pollen beetle by its effective parasitoids.
A strong negative correlation was found between pollen beetle parasitism level and the severity of cereal aphid attack, as well as between sales of the insecticide dimethoate (used to control aphids in cereals) and pollen beetle parasitism levels (Hokkanen et al. unpublished). It appears clear that in outbreak years of cereal aphids – about twice in a decade in Finland during the past several decades – the pollen beetle parasitoids are killed while the aphids are controlled with insecticide sprays. In crop rotations in Finland, spring oilseed crops are usually followed by spring cereals. Therefore, the pollen beetle parasitoids emerge in the following year from fields of spring cereals, usually exactly at the same time as when aphids colonise the fields and are controlled if needed (around mid-June till end of June). It would be best not to spray insecticides in those fields, where rapeseed was grown in the previous year, in order to maximize the biocontrol of pollen beetle by its effective parasitoids.
Simulation models (Hokkanen and Kaukoranta, unpublished) show that parasitoids are capable of completely controlling the pollen beetles at levels much below economic thresholds. This is under the assumption that the crop management does not interfere and negate their impact. Crop spraying with insecticides and soil cultivation methods are the key factors in the current management, preventing effective natural control of the beetle by its parasitoids. Both of these can easily be modified to allow effective biological control (Hokkanen et al. 1988, Hokkanen 2008).
Insecticide treatments in general in the rapeseed ecosystem appear to have led to localized eradications of the specific EPF Nosema meligethi, previously classified as Protozoa (Lipa and Hokkanen 1992, Hokkanen and Lipa 1995). Nosema meligethi is an intracellular obligate parasite specific to the genus Meligethes. It is an ideal insect pathogen from a population dynamics point of view. Nosema meligethi causes chronical disease with lowered fecundity and lifespan, and high overwintering mortality, and is transmitted both horizontally and vertically (Hokkanen and Lipa 1995). Lipa and Hokkanen (1992) studied the occurrence of N. meligethi across Europe. Samples of Meligethes spp. from 12 countries in Eastern and Western Europe were inspected for the occurrence of infection. A total of 13 910 individual beetles were checked, and of these, 561 were found to be infected (4.03%). Infection was not detectable in samples from the UK, Germany, Sweden, or Switzerland. In contrast, the disease was found rather regularly in samples from Finland and from Eastern European countries. No obvious reason for this pattern of infection is known, but pesticide usage at or close to the sampling sites may play a role. Most samples from which Nosema was detected originated from areas where few pesticides are used.
Nosema infection was also shown to lower pollen beetle weights on the average by 13% – similar to the effect of soil treatment with B. bassiana – which is enough to explain the reported increases in the overwintering mortality of the beetles due to Nosema infection (Hokkanen and Lipa 1995).
Biopesticides and entomovectoring
Only a few options are available as potential biopesticides for the control of the pollen beetle. Besides EPF and EPN, no known microbial pesticides are effective against this beetle. With EPF sprays in the field, good reduction of the pest has been obtained. For example, M. anisopliae treatment caused 75% mortality in pollen beetle larvae (Hokkanen 2008). Reduction of pollen beetle larvae on the inflorescences, however, may not be interesting from practical crop protection point of view, because the main damage to the crop has already been done by the adults. Furthermore, even a high reduction in the size of the larval population is not likely to lower the population of the new generation emerging during the summer (Hokkanen 2000). Metarhizium sprays could possibly be used to kill adult pollen beetles in the spring via fungal sprays at the bud stage. The best option might be to grow and treat an early flowering trap crop (e.g., turnip rape) with the EPF.
Targeted, precision delivery of microbiological control agents such as EPF has been developed in the last two-three decades to improve practical and economic efficiency of biocontrol agent use. Usually, managed pollinating insects such as honey bees and bumble bees are employed as vectors; hence the term ‘entomovector technology’ (first used by Hokkanen and Menzler-Hokkanen 2007). The vectoring insects are loaded with the powdery biocontrol preparate at the exit from their hive, and while visiting the flowers of the target crop they deliver the biocontrol agent onto the crop inflorescences (Hokkanen et al. 2015). As pollen beetles feed in open flowers, they are good targets for entomovectoring, because rapeseed flowers are highly attractive also to pollinators.
Butt et al. (1998) report that honey bees foraging from a hive fitted with an inoculum dispenser at the entrance effectively delivered dry conidia of M. anisopliae to the flowers of oilseed rape in caged field plots. In both winter- and spring-sown rape experiments, a greater mortality of pollen beetles occurred in treated plots than in control plots. The mortality (61% on winter rape, 100% on spring rape) was greatest during peak flowering, when the feeding activity of both bees and beetles from the flowers was maximal. This situation provided optimal conditions for inoculum dissemination and infection. Conidial sporulation occurred on a significant proportion of the dead pollen beetles.
Carreck et al. (2007) show in a later study that M. anisopliae, disseminated to the flowering canopy of oilseed rape by honey bees, infect seed weevil adults and pollen beetle adults and larvae in the flowers. Subsequent mortality of adults of both species and the larvae of pollen beetle, following incubation in the laboratory was high, and always exceeded 70%. A high proportion of cadavers showed external conidiation, confirming infection by M. anisopliae.
Experience so far shows that bees are more efficient than conventional sprayers in delivering the EPF inoculum to the pest infested flowers (Butt and Copping 2000). Entomovectoring improves targeting and provides cost-effective control of pests (see also Hokkanen et al. 2015), and can be adopted for improving pest management in rapeseed.
Entomovectoring may be most appropriate in connection with an early-flowering trap crop (Hokkanen, 1991). Field experiments have indicated excellent potential for employing Brassica rapa as a trap crop for B. napus, as it flowers clearly earlier and is highly attractive to rapeseed pests, as well as to bees and other pollinators such as hoverflies (Fig. 3) (Hokkanen 1989a).
indicated excellent potential for employing Brassica rapa as a trap crop for B. napus, as it flowers clearly earlier and is highly attractive to rapeseed pests, as well as to bees and other pollinators such as hoverflies (Fig. 3) (Hokkanen 1989a).
Conclusion
The scientific community needs to develop, design and implement integrated systems for optimised provision of ecosystem services and use of plant protection tools, with focus on ecological, economic and social sustainability of the integrated system. To facilitate full integration, leading to integrative biological control, there is a need to find ways to support biological control in agroecosystems as much as possible, and to develop “bioinspired” plant protection products and tools, based on the use of natural products active against pests and pathogens, or that trigger plant defense responses. These products and tools are needed as successor technology to replace phased-out chemical pesticides.
This is why we need to increase our knowledge on stacking of the most promising and productive ecosystem services into an operational, profitable, and sustainable whole. In the specific case of the pollen beetle, we are confident that this problem pest can be brought under complete biological control by combining several of the ecostacking component techniques into one management system.
Acknowledgements
We gratefully acknowledge the support to us from the organizers to attend the First International Congress of Biological Control, May 14-16, 2018·Beijing, China. We further acknowledge the support to our work on sustainable pest management from the EU-ERA-Net program C-IPM project “IPM4Meligethes”, from the EU-ERA-Net program LEAP-AGRI project “Pest Free Fruit”, and from the Academy of Finland project “Resilient, productive, and sustainable cropping of cassava in SE Asia: utilizing bottom up effects to manage plant-herbivore-natural enemy interactions”. Our concept of ecostacking has been influenced to some extent by colleagues participating in the preparation of a proposal to address the EU-call SFS-28-2017: Functional biodiversity – productivity gains through functional biodiversity: effective interplay of crop pollinators and pest predators.
References
Balzan MV (2017) Flowering banker plants for the delivery of multiple agroecosystem services. Arthropod Plant Interact 11:743, https://doi.org/10.1007/s11829-017-9544-2
Brandes M, Heimbach U, Ulber B (2018) Effects of insecticide application on parasitism rates of pollen beetle larvae (Brassicogethes aeneus (Fabricius)) by tersilochine parasitoids. Arthropod-Plant Interactions, https://doi.org/10.1007/s11829-017-9580-y
Butt TM, Carreck NL, Ibrahim L, Williams IH (1998) Honey-bee-mediated Infection of pollen beetle (Meligethes aeneus Fab.) by the insect-pathogenic fungus, Metarhizium anisopliae. Biocontrol Science and Technology 8: 533-538, http://dx.doi.org/10.1080/09583159830045
Butt TM, Copping LG (2000) Fungal biological control agents. Pesticide Outlook 11, 186-191, http://dx.doi.org/10.1039/B008009H
Butt TM, Ibrahim L, Ball BV, Clark SJ (1994) Pathogenicity of the entomogenous fungi Metarhizium anisopliae and Beauveria bassiana against crucifer pests and the honey bee. Biocontrol Science & Technology, 4: 207-214
Carreck NL, Butt TM, Clark SJ, Ibrahim L, Isger EA, Pell JK, Williams IH (2007) Honey bees can disseminate a microbial control agent to more than one inflorescence pest of oilseed rape. Biocontrol Sci Technol 17, 179–191, http://dx.doi.org/10.1080/09583150600937485
Ekbom B, Borg A (1996) Pollen beetle (Meligethes aeneus) oviposition and feeding preference on different host plant species. Entomol Exp Appl 78:291–299
Grettenberger IM, Tooker JF (2017) Variety mixtures of wheat influence aphid populations and attract an aphid predator. Arthropod Plant Interact 11:133, https://doi.org/10.1007/s1182 9-016-9477-1
Hatt S, Uyttenbroeck R, Lopes T, Chen JL, Piqueray J, Monty A, Francis F (2018) Effect of flower traits and hosts on the abundance of parasitoids in perennial multiple species wildflower strips sown within oilseed rape (Brassica napus) crops. Arthropod-Plant Interactions https://doi.org/10.1007/s11829-017-9567-8
Hervé MR, Cortesero AM (2016) Potential for oilseed rape resistance in pollen beetle control. Arthropod Plant Interact 10: 463–475, https://doi.org/10.1007/s11829-016-9438-8
Hervé MR, Delourme R, Leclair M, Marnet N, Cortesero AM (2014) How oilseed rape (Brassica napus) genotype influences pollen beetle (Meligethes aeneus) oviposition. Arthropod Plant Interact 8: 383–392, https://doi.org/10.1007/s11829-014-9321-4
Hokkanen H, Granlund H, Husberg G-B, Markkula M (1986) Trap crops used successfully to control Meligethes aeneus (Col., Nitidulidae), the rape blossom beetle. Annales Entomologici Fennici 52: 115-120
Hokkanen H, Husberg G-B, Söderblom M (1988) Natural enemy conservation for the integrated control of the rape blossom beetle Meligethes aeneus F. Annales Agriculturae Fenniae 27: 281-293
Hokkanen HMT & Menzler-Hokkanen I (2007) Use of honeybees in the biological control of plant diseases. Entomol Res 37 (Suppl. 1), A62-A63. DOI: 10.1111/j.1748-5967.2007.00082.x
Hokkanen HMT (1989) Biological and biotechnical control of the rape blossom beetle Meligethes aeneus F. Acta Entomologica Fennica 53: 25-29
Hokkanen HMT (1991) Trap cropping in pest management. Annu Rev Entomol 36:119–138. https://doi.org/10.1146/annurev.en.36.010191.001003
Hokkanen HMT (1993) Overwintering survival and spring emergence in Meligethes aeneus: effects of body weight, crowding, and soil treatment with Beauveria bassiana. Entomologia experimentalis et applicata 67: 241-246
Hokkanen HMT (2000) The making of a pest: recruitment of Meligethes aeneus onto oilseed brassicas. Entomologia Experimentalis et Applicata 95: 141-149
Hokkanen HMT (2005) Impact of predators on pollen beetle Meligethes aeneus on rapeseed in Finland. IOBC/WPRS Bulletin 27: 295-298
Hokkanen HMT (2006) Phradis morionellus on Meligethes aeneus: long-term patterns of parasitism and impact on pollen beetle populations in Finland. IOBC/wprs Bulletin 29(7): 187-192
Hokkanen HMT (2008) Biological control methods of pest insects in oilseed rape. Bulletin OEPP/EPPO Bulletin 38: 104–109
Hokkanen HMT (2015) Integrated pest management at the crossroads: Science, politics, or business (as usual)? Arthropod-Plant Interactions 9: 543-545, http://dx.doi.org/10.1007/s11829-015-9403-y
Hokkanen HMT (2017) Ecostacking: maximising the benefits of ecosystem services. Arthropod-Plant Interactions 11: 741, https://doi.org/10.1007/s11829-017-9575-8
Hokkanen HMT, Lipa JJ (1995) Occurrence and dynamics of Nosema meligethi (Microsporida) in populations of Meligethes aeneus (Coleopera, Nitidulidae) in Finland. Entomologica Fennica 6: 11-18
Hokkanen HMT, Menzler-Hokkanen I (2007) Use of honeybees in the biological control of plant diseases. Entomol Res 37 (Suppl. 1), A62-A63, https://doi.org/10.1111/j.1748-5967.2007.00082.x
Hokkanen HMT, Menzler-Hokkanen I (2017) The use of entomopathogenic fungi in the insect pest management of Brassica oilseed crops. In: Reddy GVP (ed) Integrated management of insect pests on canola and other Brassica oilseed crops. CABI, Boston, pp 373–382
Hokkanen HMT, Menzler-Hokkanen I (2017) Use of entomopathogenic fungi in the insect pest management of Brassica oilseed crops. In: Reddy GVP (ed) Integrated management of insect pests on canola and other Brassica oilseed crops. CAB International, Wallingford, pp 373–382
Hokkanen HMT, Menzler-Hokkanen I (2018a) Insect pest suppressive soils: buffering pulse cropping systems against outbreaks of Sitona weevils. Ann Entomol Soc Am. https://doi.org/10.1093/aesa/say019
Hokkanen HMT, Menzler‑Hokkanen I (2018b) Urgent need to develop ecostacking techniques to enhance ecosystem services in cropping systems. Arthropod-Plant Interactions https://doi.org/10.1007/s11829-018-9620-2
Hokkanen HMT, Menzler‑Hokkanen I (2018c) Trap cropping as a delivery platform for RNAi-based targeted control of the pollen beetle Brassicogethes aeneus. Mitt. Dtsch. Ges. allg. angew. Ent. 21 (in press).
Hokkanen HMT, Menzler‑Hokkanen I, Butt TM (2003) Pathogens of oilseed rape pests. In: Alford DV (Ed.) Biocontrol of Oilseed Rape Pests. Blackwell Science, Oxford, UK, pp. 299-322
Hokkanen HMT, Menzler-Hokkanen I, Lahdenperä M-L ( 2015) Managing Bees for Delivering Biological Control Agents and Improved Pollination in Berry and Fruit Cultivation. Sustainable Agriculture Research 4, No. 3; 2015, https://dx.doi.org/10.5539/sar.v4n3p89
Hornby D (1983) Suppressive Soils. Annual Review of Phytopathology 21:65-85, https://doi.org/10.1146/annurev.py.21.090183.000433
Husberg G-B, Hokkanen HMT (2001) Efects of Metarhizium anisopliae on the pollen beetle Meligethes aeneus and its parasitoids Phradis morionellus and Diospilus capito. BioControl 46: 261-273
Lipa JJ, Hokkanen HMT (1992) Nosema meligethi I. & R. (Microsporida) in populations of Meligethes spp. in Europe. Biocontrol Science and Technology 2: 119-125
Mauchline AL, Cook SM, Powell W, Chapman JW, Osborne JL (2017) Migratory flight behaviour of the pollen beetle Meligethes aeneus. Pest Management Science, 73: 1076–1082, https://doi.org/10.1002/ps.4550
Menzler-Hokkanen I, Hokkanen HMT (2005) Developing entomopathogenic nematode delivery systems for biological control of oilseed rape pests. IOBC/WPRS Bulletin 28: 19-22
Menzler-Hokkanen I, Hokkanen HMT (2018) Socioeconomics of adopting IPM strategies by oilseed rape farmers. EPPO Bulletin 48, https://doi.org/10.1111/epp.12498
Menzler-Hokkanen I; Hokkanen H; Büchs W; Klukowski K; Luik A; Nilsson C; Ulber B; Williams I. (2006). Insect pest problems in oilseed rape and how to deal with them: the farmers’ perspective. IOBC/wprs Bulletin 29(7): 91-94.
Müller HJ (1941) Weitere Beiträge zur Biologie des Rapsglanzkäfers Meligethes aeneus F. (Ûber das Winterlager und die Massenbewegung im Frühjahr). Z. Pflkrankh. Pflschutz 51: 529-595.
Perälä TH (1995). Rapsikuoriainen ja sen luontaiset viholliset rypsi-apila sekakasvustossa. Pollen beetle and its natural enemies in rapeseed-clover intercrop. MSc-Thesis, Department of Applied Zoology, University of Helsinki, Finland, 76 p.
Pineda A, Kaplan I, Bezemer TM (2017) Steering soil microbiomes to suppress aboveground insect pests. Trends Plant Sci 22:770–778, https://doi.org/10.1016/j.tplan ts.2017.07.002
Prieto JD, Castañé C, Calvet C, Camprubi A, Battaglia D, Trotta V, Fanti P (2017) Tomato belowground–aboveground interactions: Rhizophagus irregularis affects foraging behavior and life history traits of the predator Macrolophus pygmaeus (Hemiptera: Miridae). Arthropod Plant Interact 11: 15–22, https://doi.org/10.1007/s1182 9-016-9465-5
Roberts DW, St Leger RJ (2004) Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Advances in Applied Microbiology 54, 1-70, https://dx.doi.org/10.1016/S0065-2164(04)54001-7
Schlatter D, Kinkel LL, Thomashow LS, Weller DM, Paulitz TC (2017) Disease suppressive soils: new insights from the soil microbiome. Phytopathol. https://doi.org/10.1094/PHYTO-03-17-0111-RVW
Schlatter D, Kinkel LL, Thomashow LS, Weller DM, Paulitz TC (2017) Disease suppressive soils: new insights from the soil microbiome. Phytopathology, https://doi.org/10.1094/PHYTO-03-17-0111-RVW
Schröder ML, Glinwood R, Ignell R, Krüger K (2017) The role of visual and olfactory plant cues in aphid behaviour and the development of non-persistent virus management strategies. Arthropod Plant Interact 11: 1–13, https://doi.org/10.1007/s11829-016-9463-7
Shelton AM, Badenes-Perez FR (2006) Concept and Applications of Trap Cropping in Pest Management. Annual Review of Entomology, 51, 285-308, http://dx.doi.org/10.1146/annurev.ento.51.110104.150959
Shikano I, Rosa C, Tan C-W, Felton GW (2017) Tritrophic interactions: microbe-mediated plant effects on insect herbivores. Ann Rev Phytopathol 55:313–331, https://doi.org/10.1146/annurevphyto-08051 6-035319
Shrestha B, Finke D, Pinero J (2019). The ‘Botanical Triad’: The Presence of Insectary Plants Enhances Natural Enemy Abundance on Trap Crop Plants in an Organic Cabbage Agro-Ecosystem. Insects 10: 181. https://doi.org/10.3390/insects10060181
Skellern MP, Cook SM (2018) Prospects for improved off-crop habitat management for pollen beetle control in oilseed rape. Arthropod-Plant Interactions, https://doi.org/10.1007/s11829-018-9598-9
St Leger RJ, Allee LL, May B, Staples RC, Roberts DW (1992) World-wide distribution of genetic variation among isolates of Beauveria spp. Mycological Research 96, 1007-1015, http://dx.doi.org/10.1016/S0953-7562(09)80108-1
Taimr L, Sedivy J, Bergmannova E and Hanker I (1967) Further experience obtained in studies on dispersal flights of Meligethes aeneus F., marked with P32 (Coleoptera). Eur J Entomol 64:325–332
Thies C, Tscharntke T (1999) Landscape structure and biological control in agroecosystems. Science 285:893–895, https://doi.org/10.1126/science.285.5429.893
Tiilikainen TM, Hokkanen HMT (2008) Pyrethroid resistance in Finnish pollen beetle (Meligethes aeneus) populations – is it around the corner? Bulletin OEPP/EPPO Bulletin 38: 99–103
Vänninen I (1996) Distribution and occurrence of four entomopathogenic fungi in Finland: effect of geographical location, habitat type and soil type. Mycological Research 100, 93-101
Vänninen I, Husberg G-B, Hokkanen H (1989) Occurrence of entomopathogenic fungi and entomophilic nematodes in cultivated soils in Finland. Acta Entomologica Fennica 53: 65-71
Veromann E, Saarniit M, Kevvai R, Luik A (2009) Effect of crop management on the incidence of Meligethes aeneus Fab. and their larval parasitism rate in organic and conventional winter oilseed rape. Agron Res 7:548–554
